Population genetics of rapid adaptation and the predictability of evolution
Richard Neher
Biozentrum, University of Basel
slides at neherlab.org/201704_UBasel_Zoology.html
Evolution of HIV
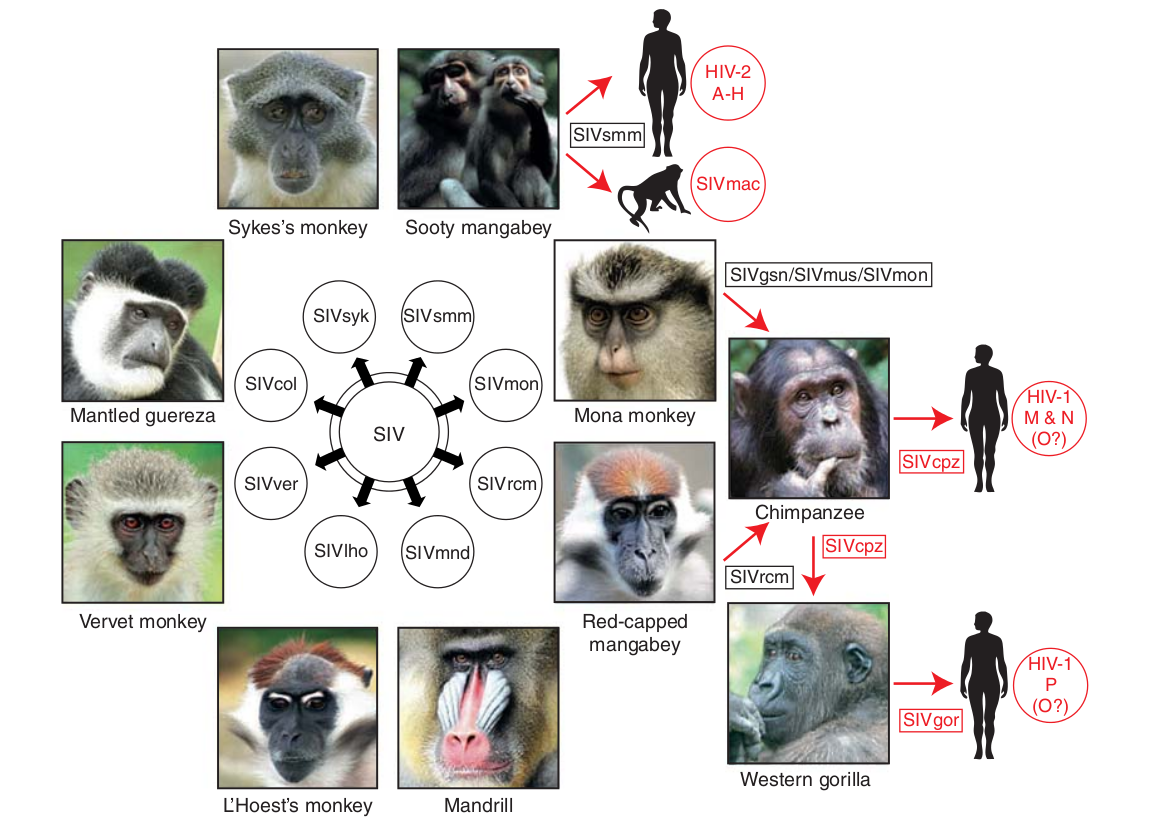
- Chimp → human transmission ~1900 gave rise to HIV-1 group M
- Diversified into subtypes that are ~20% different
- evolves at a rate of about 0.1% per year
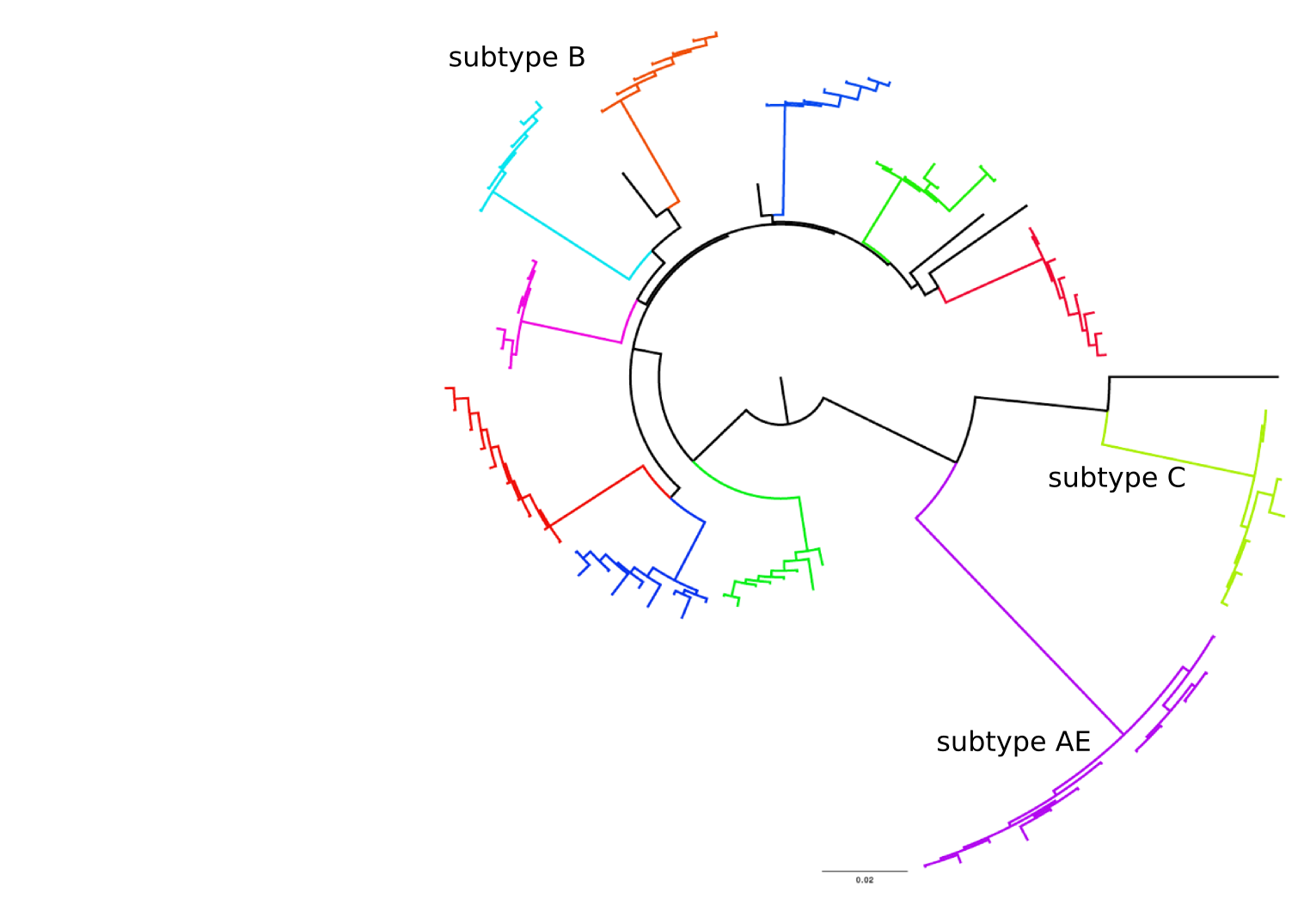
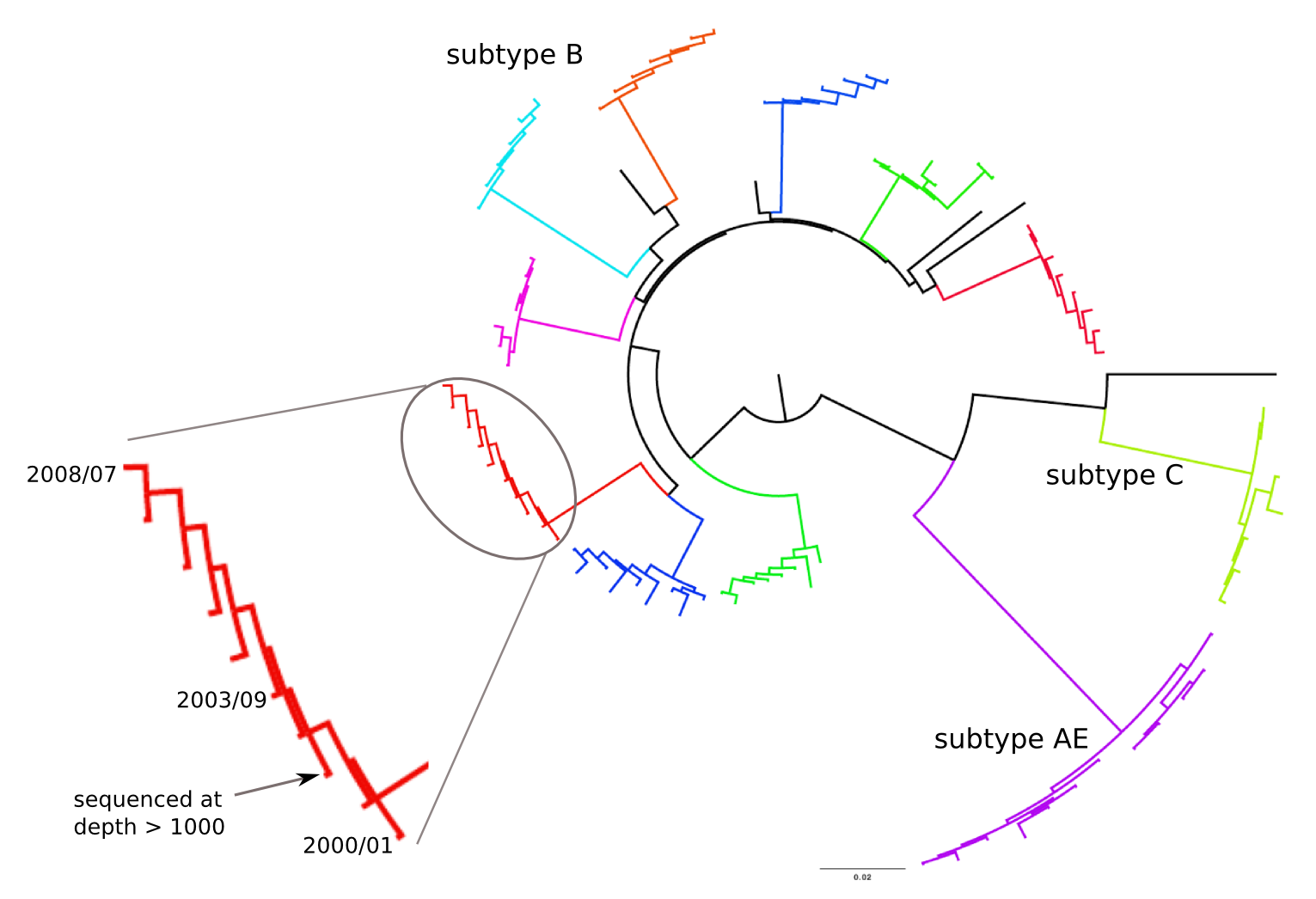
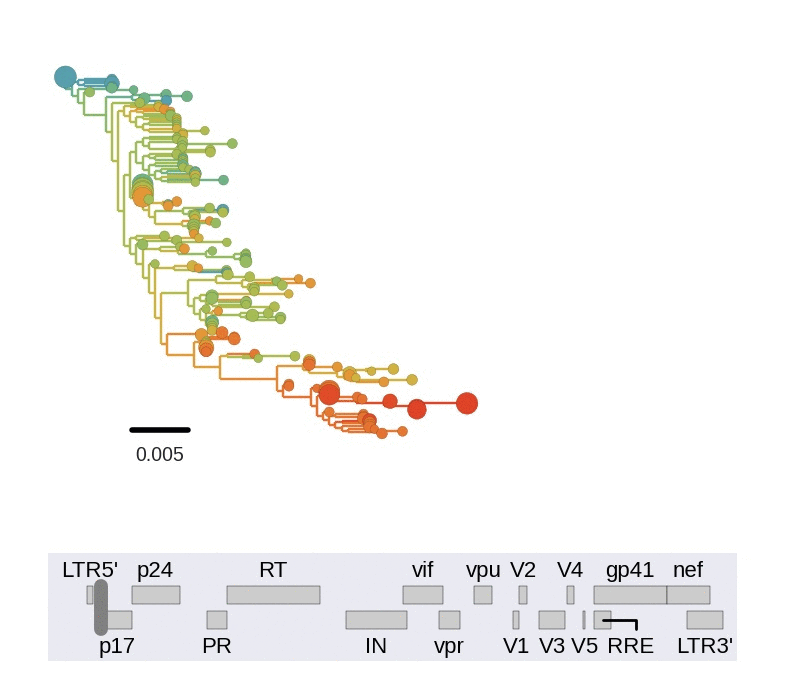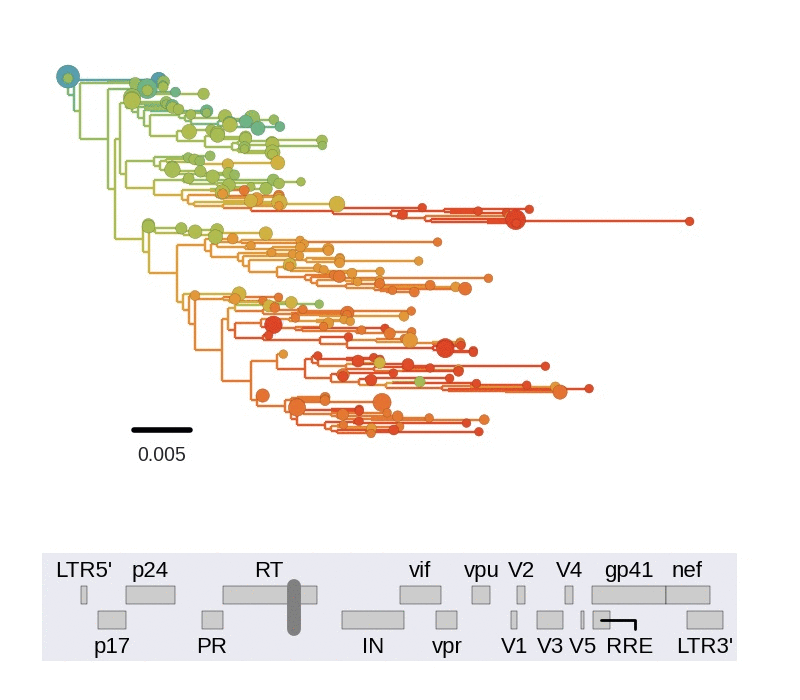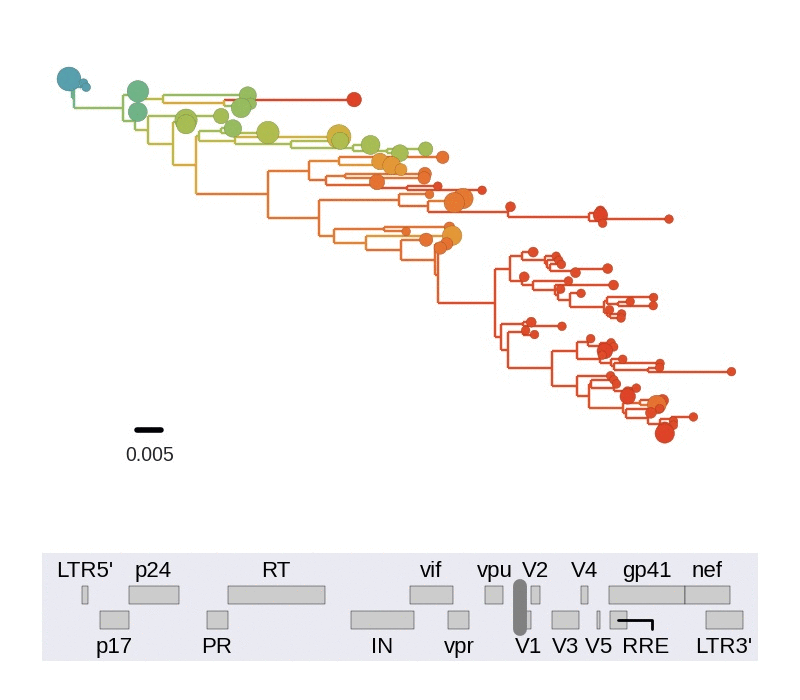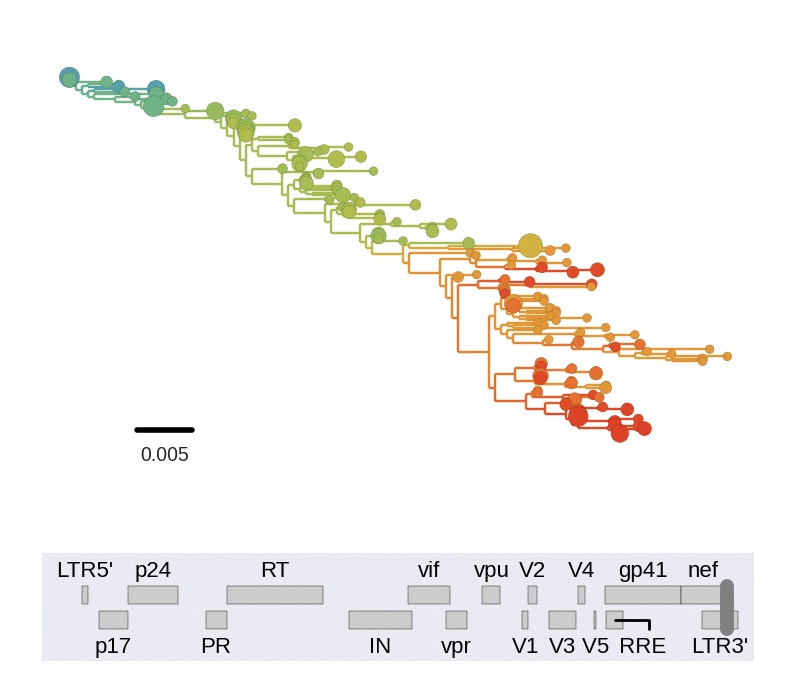
Frequent version of previously beneficial mutations
- HIV escapes immune systems
- most mutations are costly
- humans selects for different mutations
- compensation or reversion?
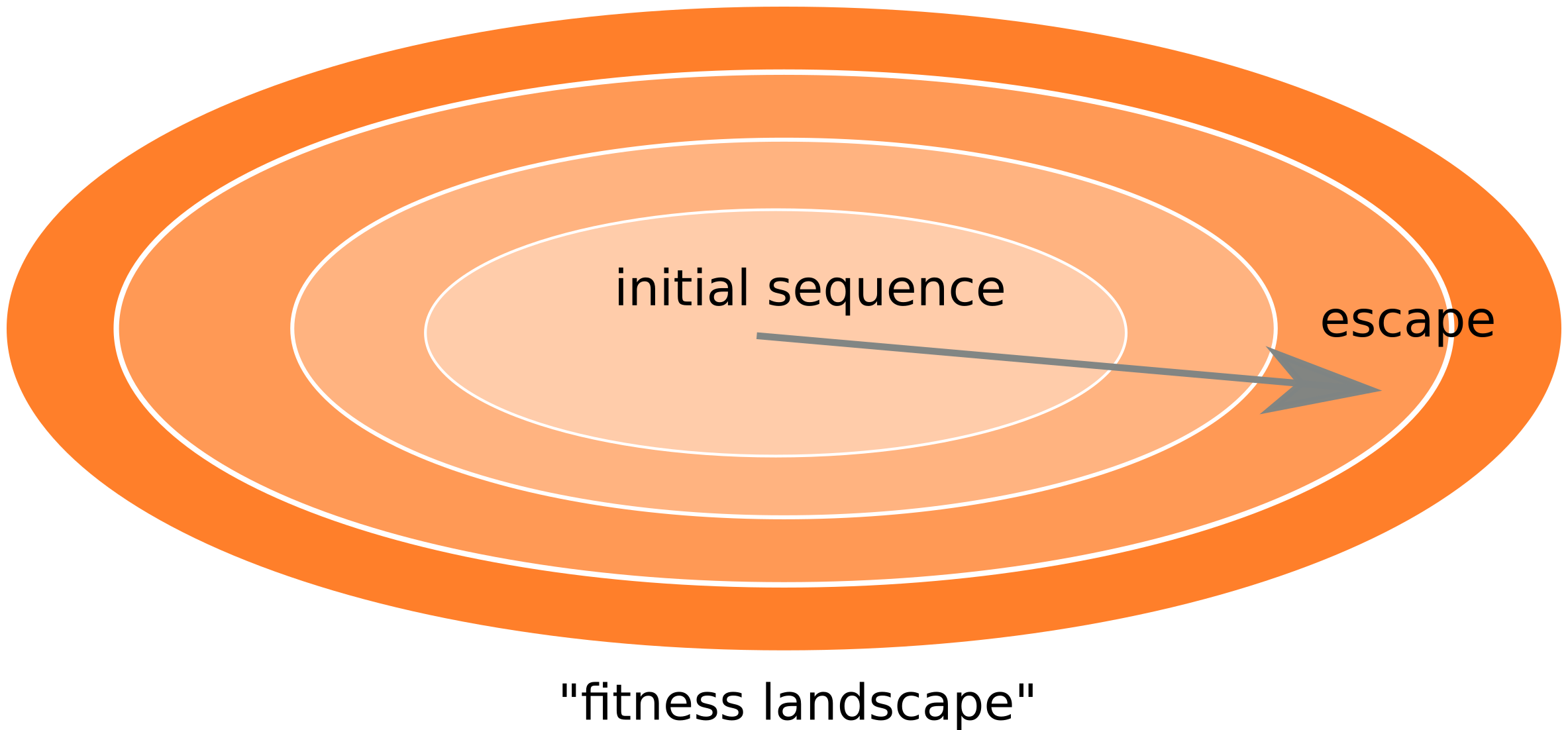
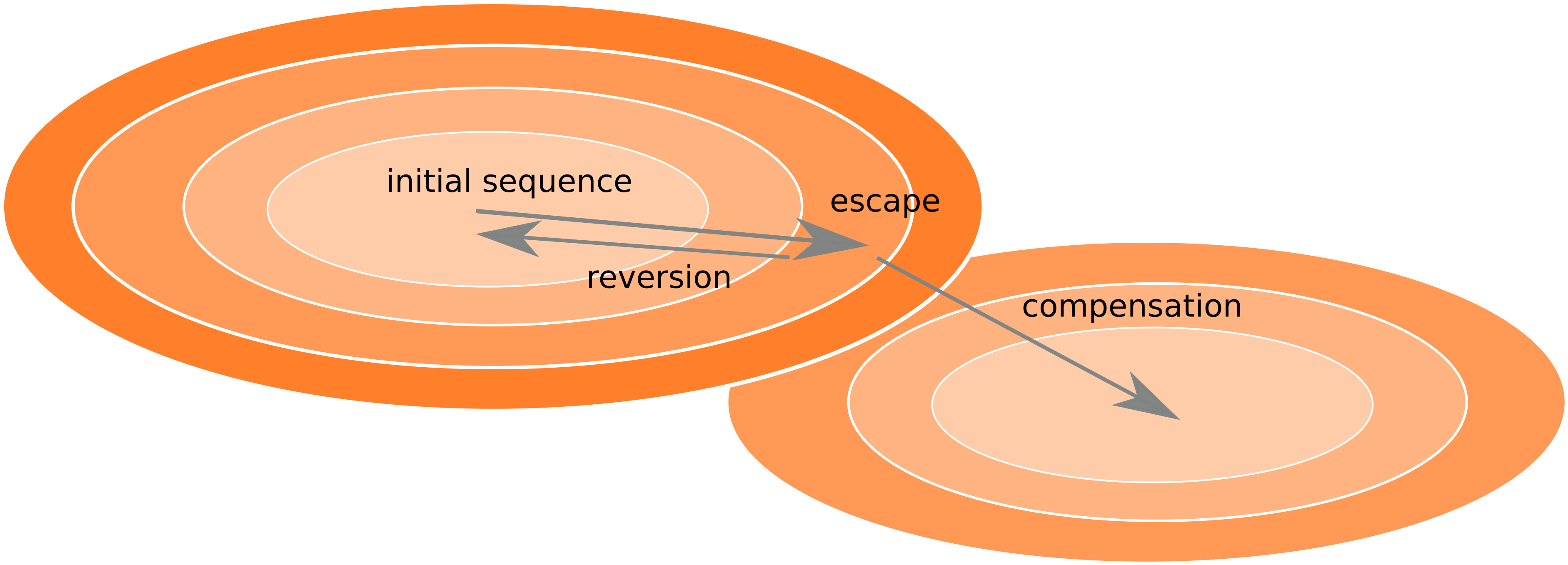
Inference of fitness costs
- mutation away from preferred state with rate $\mu$
- selection against non-preferred state with strength $s$
- variant frequency dynamics: $\frac{d x}{dt} = \mu -s x $
- equilibrium frequency: $\bar{x} = \mu/s $
- fitness cost: $s = \mu/\bar{x}$
Fitness landscape of HIV-1
Zanini et al, Virus Evolution, 2017Selection on RNA structures and regulatory sites
Zanini et al, Virus Evolution, 2017The distribution of fitness costs
Zanini et al, Virus Evolution, 2017Fitness - diversity correlation
Zanini et al, Virus Evolution, 2017Clonal interference and traveling waves
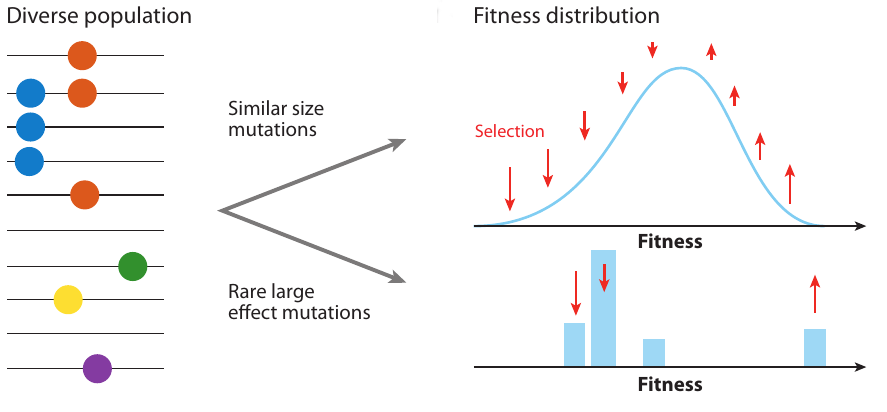
- extensive work on speed of adaptation, but this speed is not observable
- genetic diversity is what we observe
- depends on the properties of trees
What if selection is everywhere?
Kingman coalescent
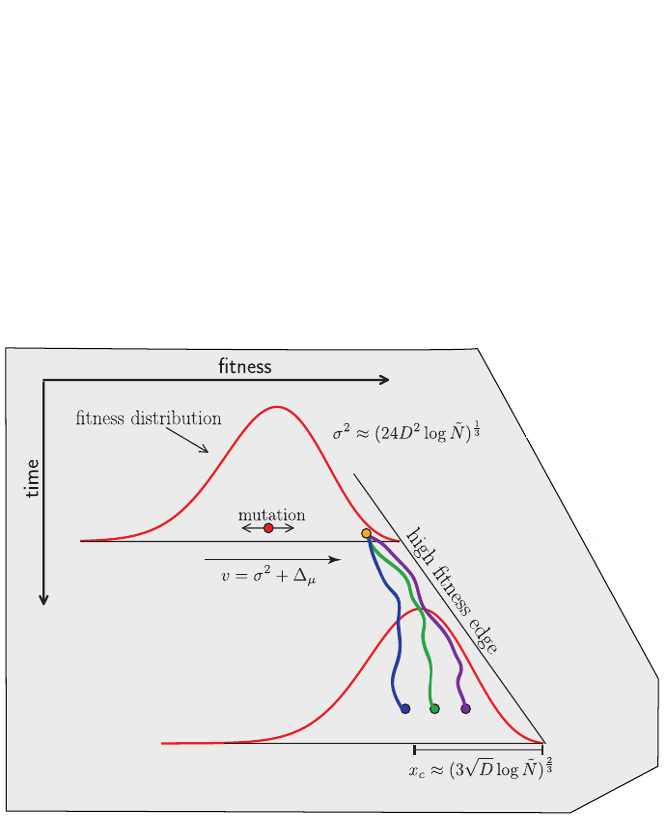
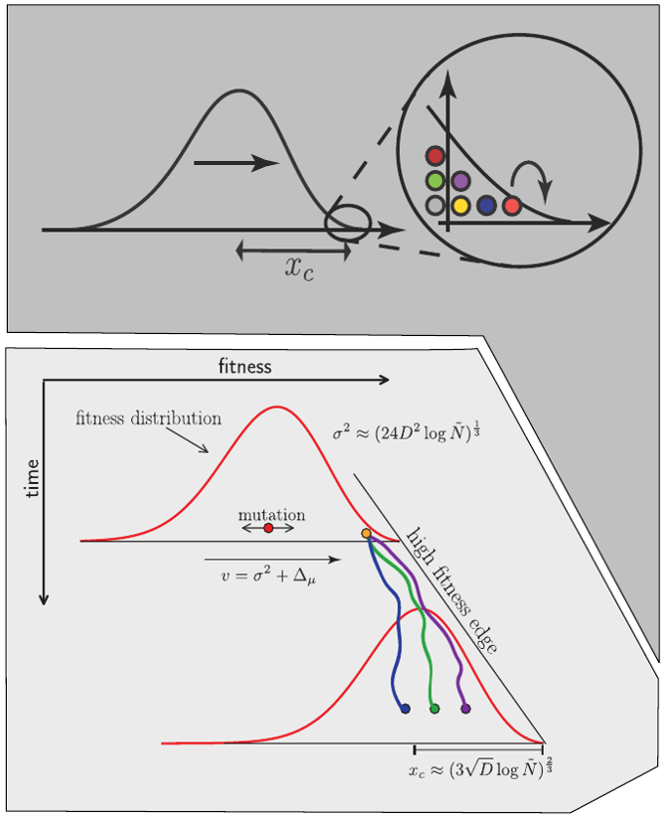
strong selection
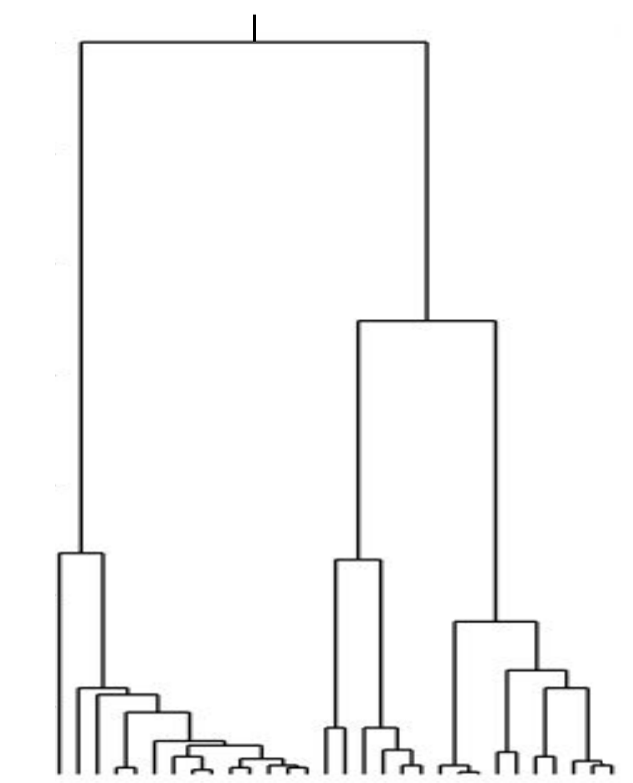
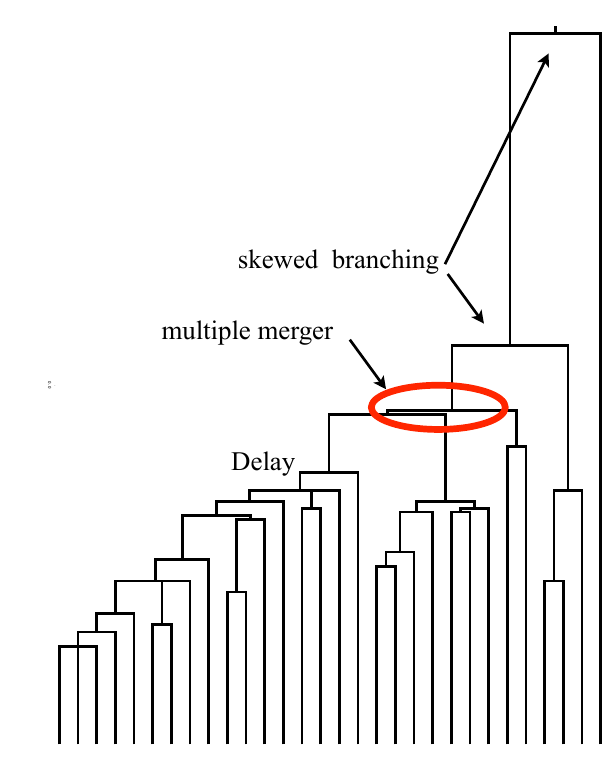
Bolthausen-Sznitman Coalescent
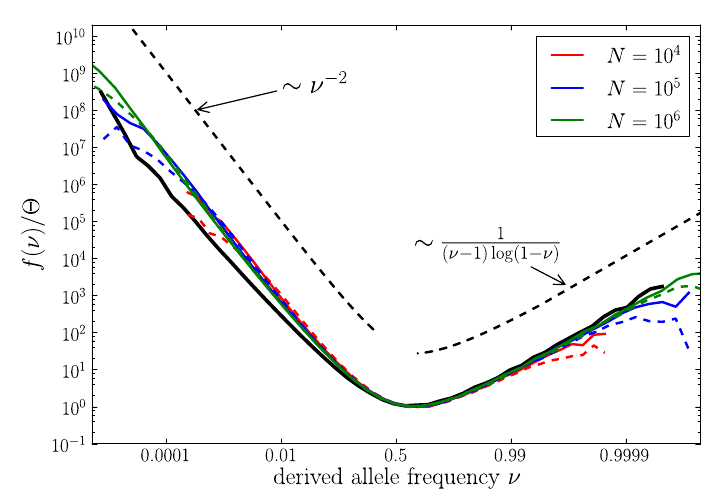
U-shaped polarized site frequency spectra

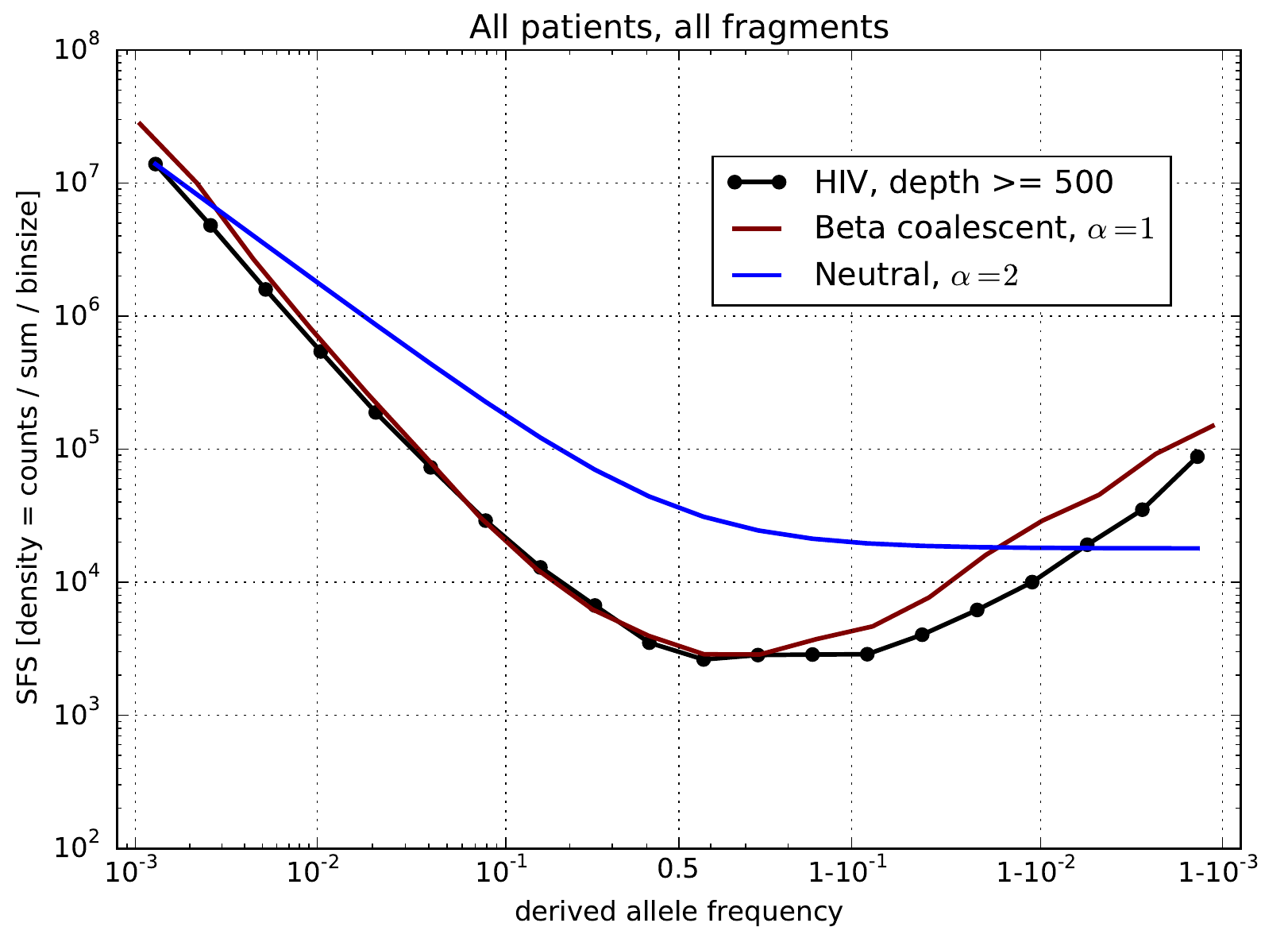 Zanini et al, eLife, 2015
Zanini et al, eLife, 2015
Bursts in a tree ↔ high fitness genotypes
Can we read fitness of a tree?
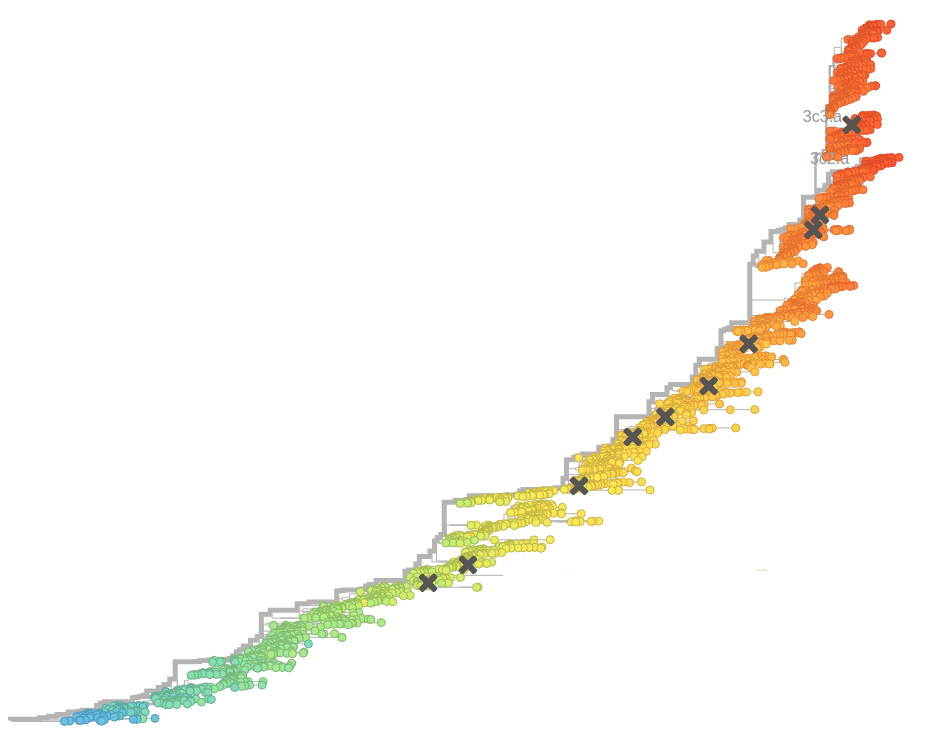
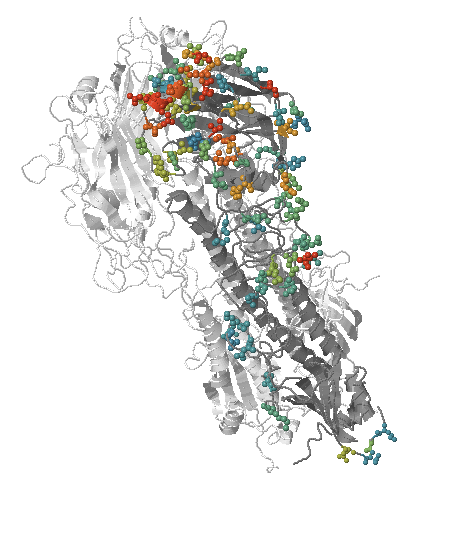
- Influenza virus evolves to avoid human immunity
- Vaccines need frequent updates
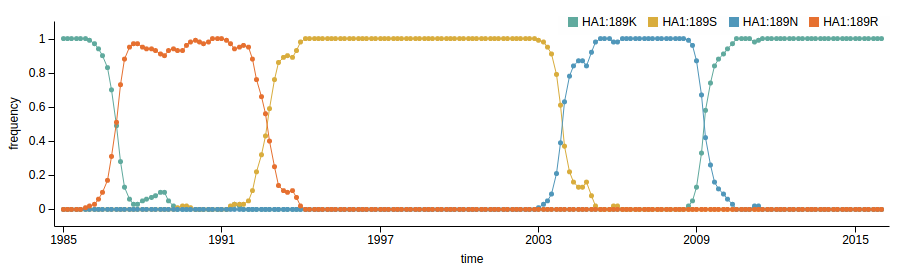
nextflu.org
nextstrain.org
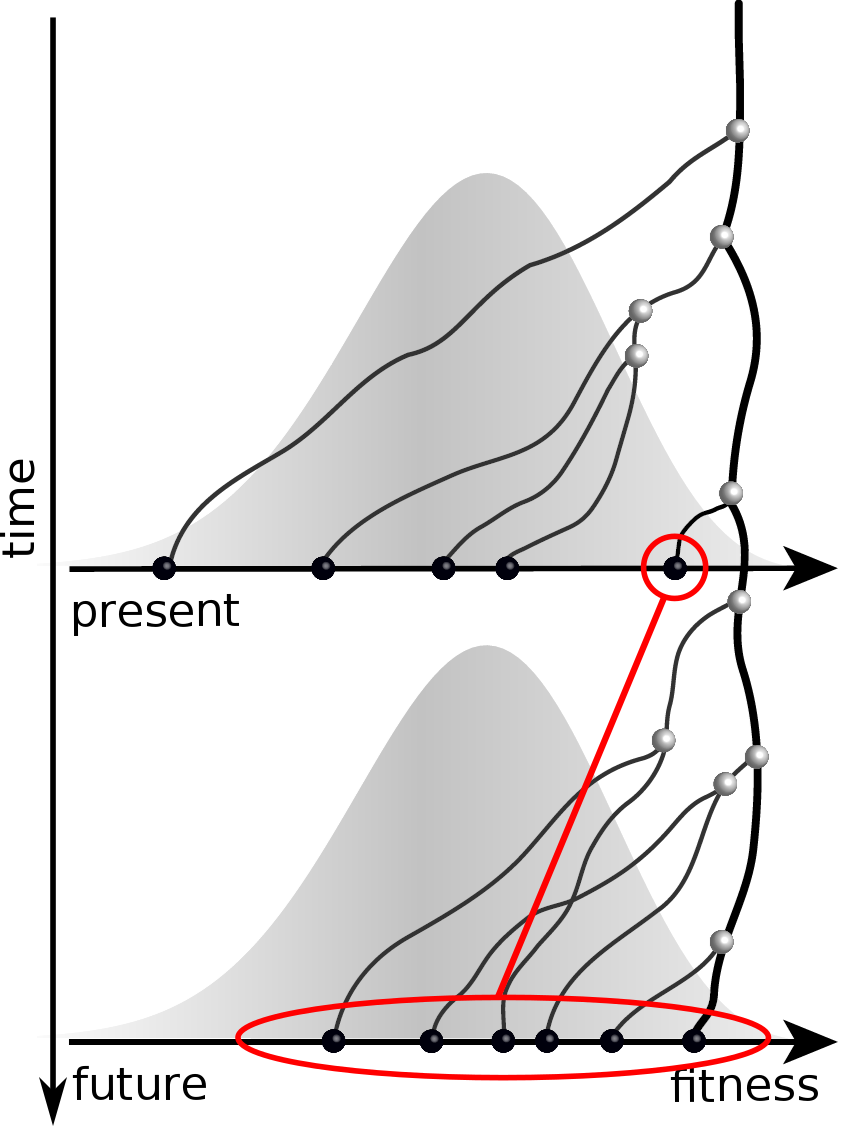
Predicting evolution
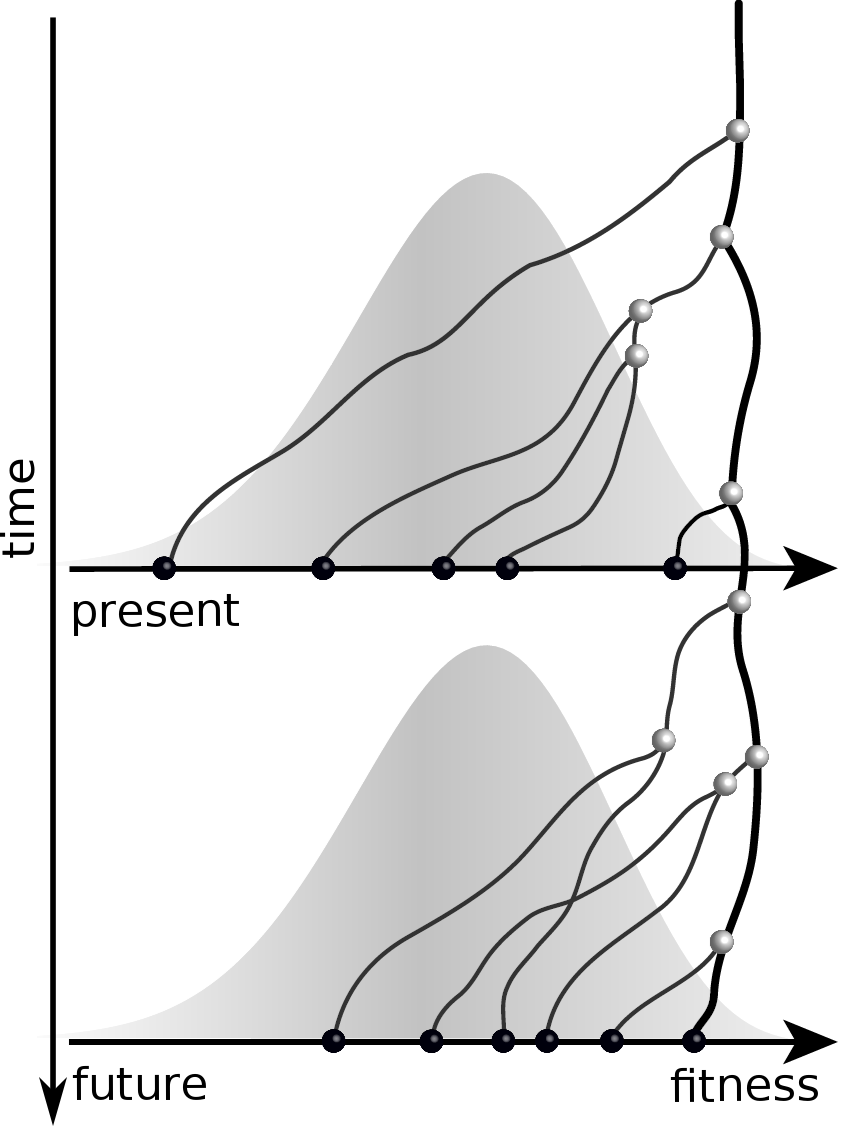
Given the branching pattern,
RN, Russell, Shraiman, eLife, 2014
- can we predict fitness?
- pick the closest relative of the future?
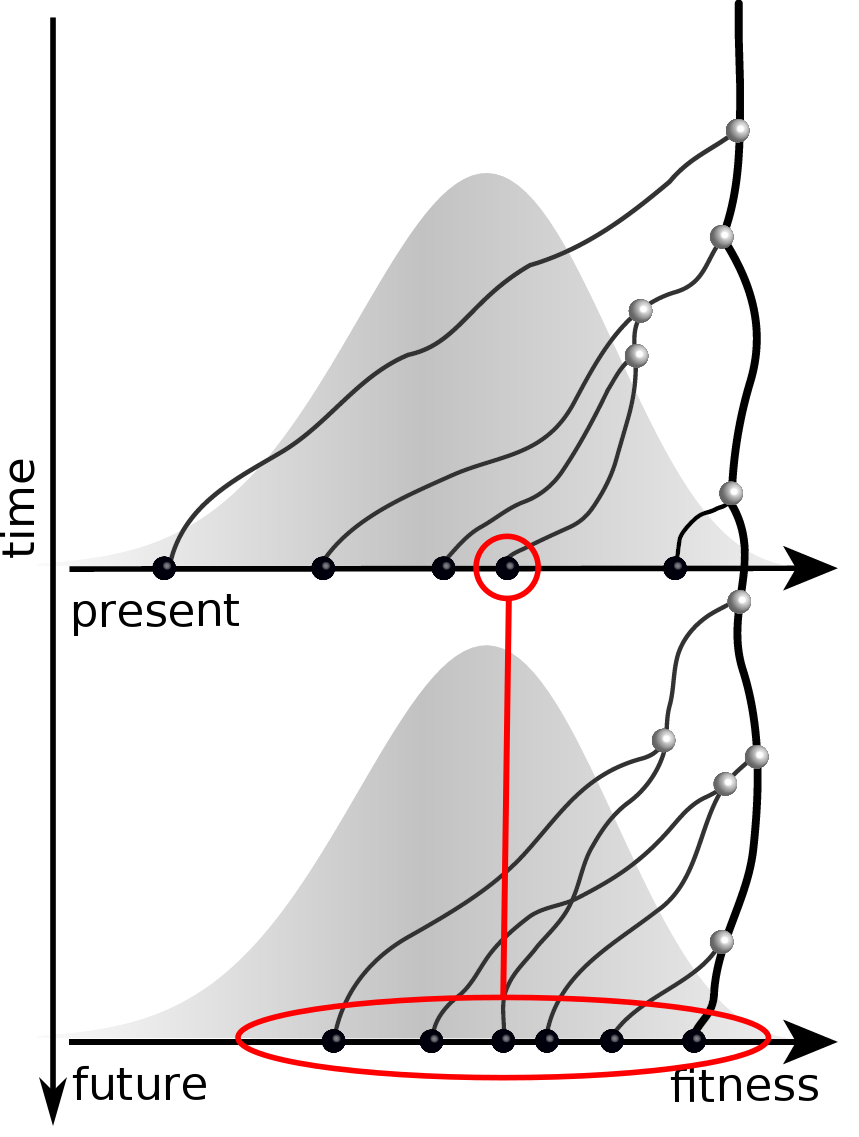
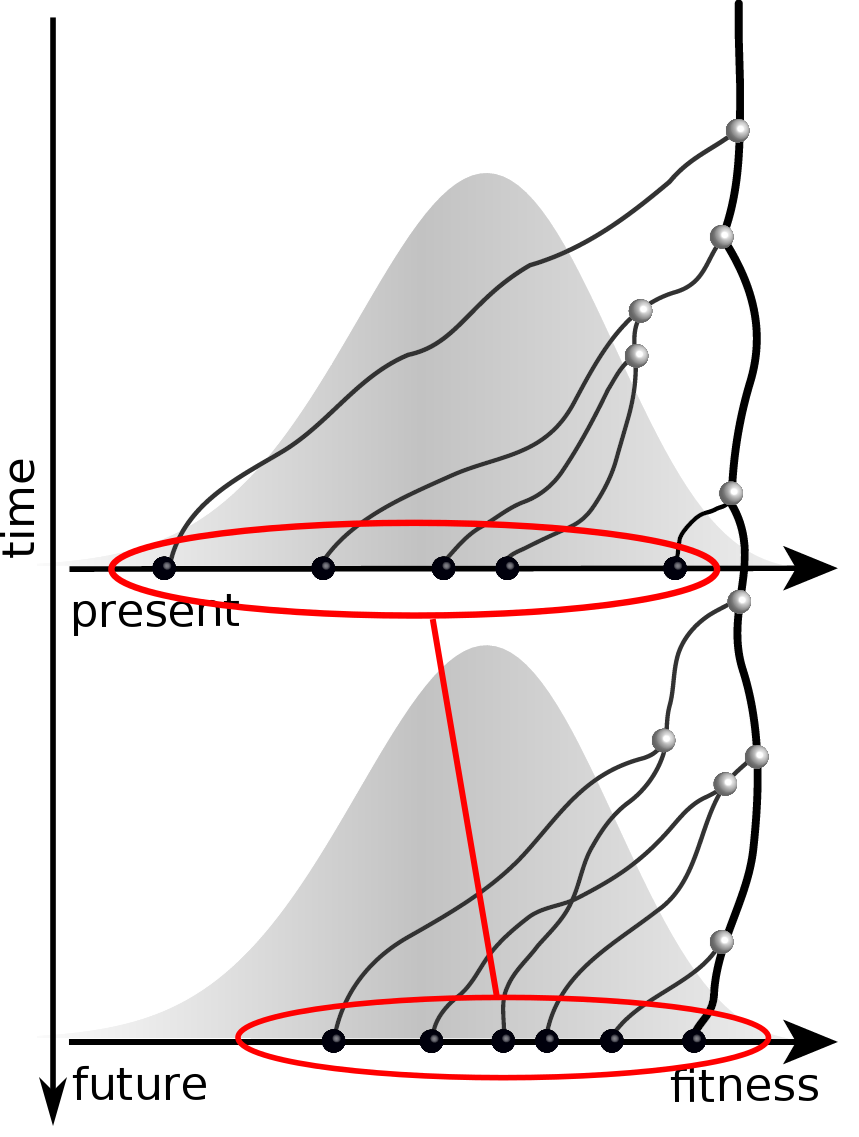
Validate on simulation data
- simulate evolution
- sample sequences
- reconstruct trees
- infer fitness
- predict ancestor of future
- compare to truth
Validation on simulated data
Prediction of the dominating H3N2 influenza strain
RN, Russell, Shraiman, eLife, 2014HIV acknowledgments
- Fabio Zanini
- Jan Albert
- Johanna Brodin
- Christa Lanz
- Göran Bratt
- Lina Thebo
- Vadim Puller
Influenza and Theory acknowledgments




- Boris Shraiman
- Colin Russell
- Trevor Bedford
- Oskar Hallatschek