The evolution of untreated HIV and maintenance of the latent reservoir
Richard Neher
Biozentrum, University of Basel
slides at neherlab.org/201809_PEI.html
Evolution of HIV
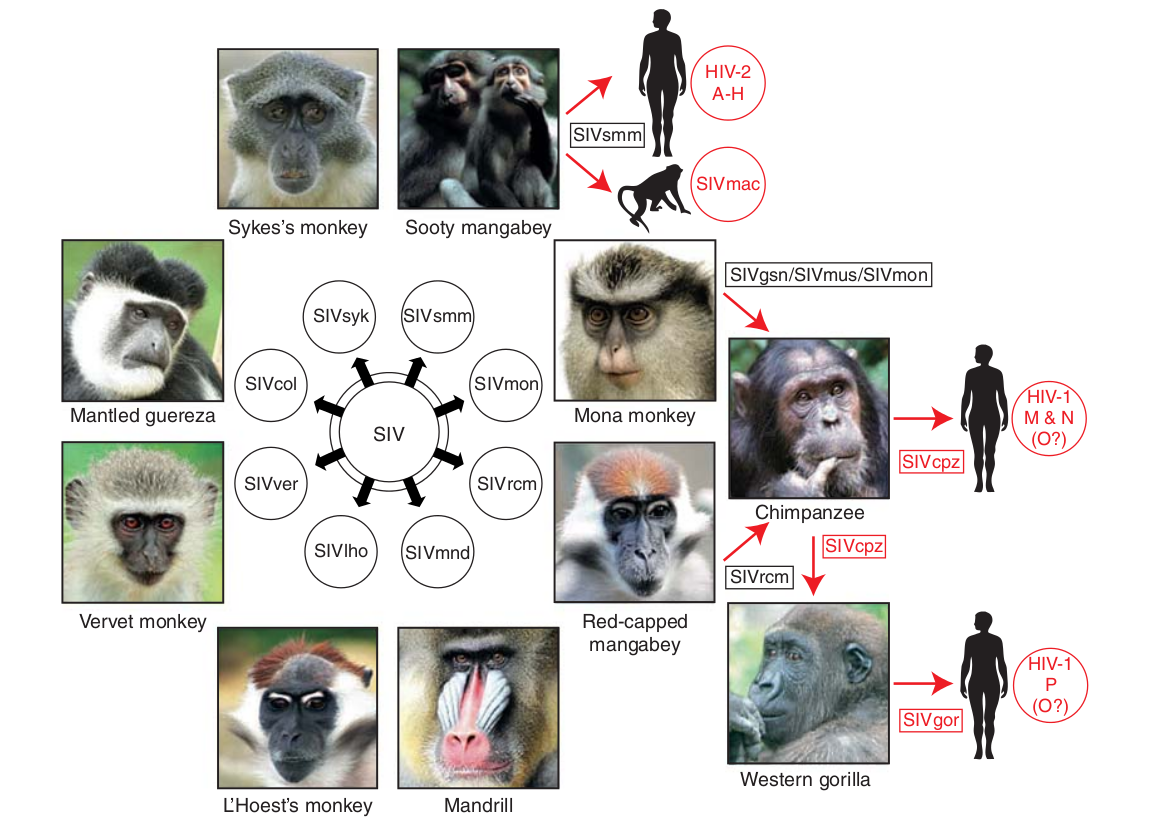
- Chimp → human transmission around 1900 gave rise to HIV-1 group M
- ~100 million infected people since
- subtypes differ at 10-20% of their genome
- HIV-1 evolves ~0.1% per year
HIV infection
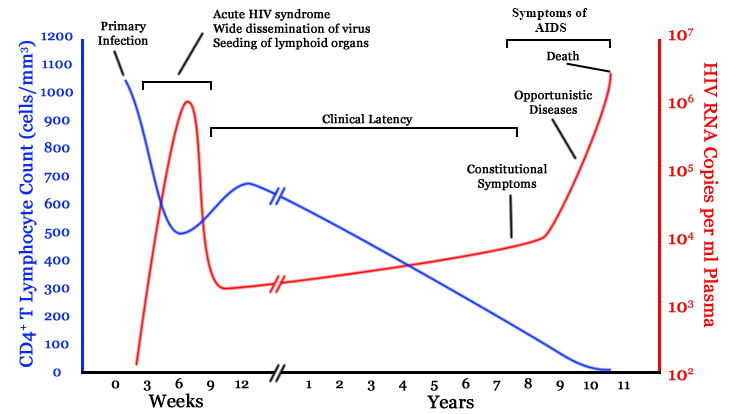

- $10^8$ cells are infected every day
- the virus repeatedly escapes immune recognition
- integrates into T-cells as
latent provirus
{kind=link}
HIV-1 evolution within one individual

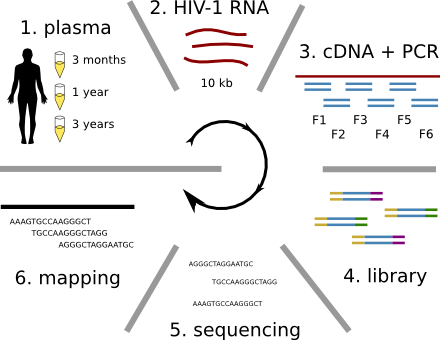

HIV-1 sequencing before and after therapy
 Zanini et al, eLife, 2015;
Brodin et al, eLife, 2016.
Collaboration with the group of Jan Albert
Zanini et al, eLife, 2015;
Brodin et al, eLife, 2016.
Collaboration with the group of Jan Albert
Approximately neutral divergence -- silent mutations
 Zanini et al, Virus Evolution, 2017
Zanini et al, Virus Evolution, 2017
In vivo mutation rate estimates
 Zanini et al, Virus Evolution, 2017
Zanini et al, Virus Evolution, 2017
Divergence at increasingly conserved positions
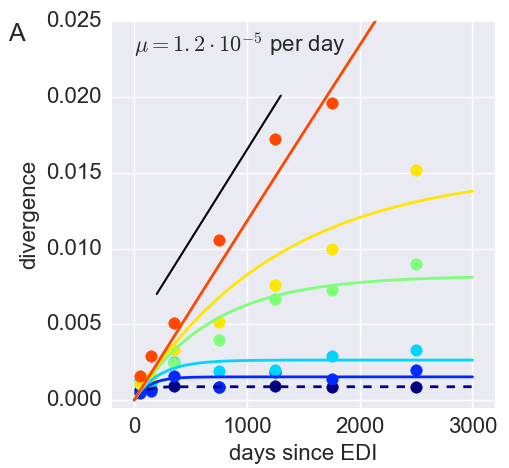
- Six categories from high to low conservation
- mutation away from preferred state with rate $\mu$
- selection against non-preferred state with strength $s$
- variant frequency dynamics: $\frac{d x}{dt} = \mu -s x $
- equilibrium frequency: $\bar{x} = \mu/s $
- fitness cost: $s = \mu/\bar{x}$
- Fit model of minor variation to categories of conservation
- $\Rightarrow$ harmonic average fitness cost in category
Accurate frequency estimates by averaging many samples

- Frequencies of costly mutations decorrelate fast $\frac{d x}{dt} = \mu -s x $
- $\Rightarrow$ average many samples to obtain accurate estimates
Fitness landscape of HIV-1
Zanini et al, Virus Evolution, 2017Selection on RNA structures and regulatory sites
- Blue: all mutations
- Red: only mutations that don't change amino acids
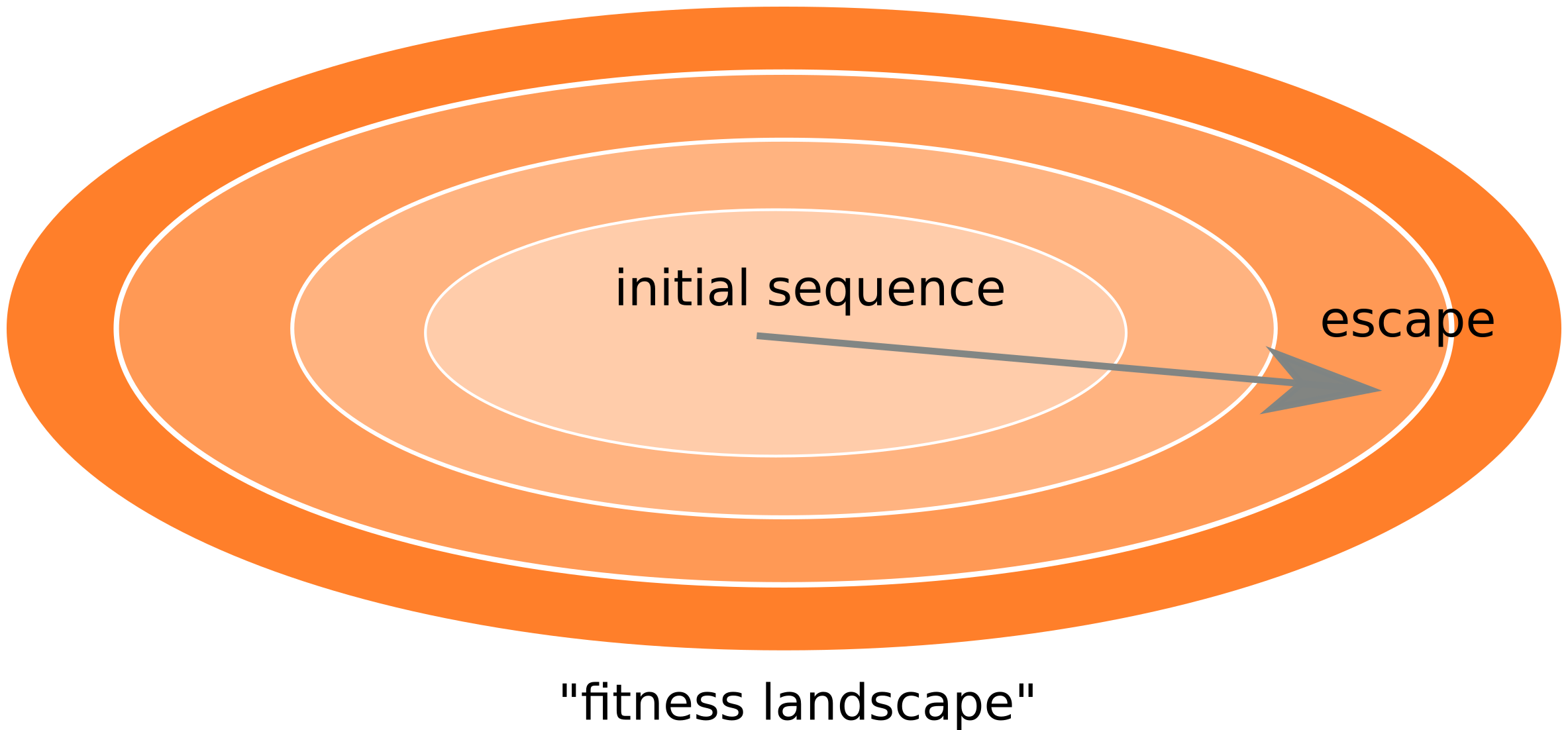
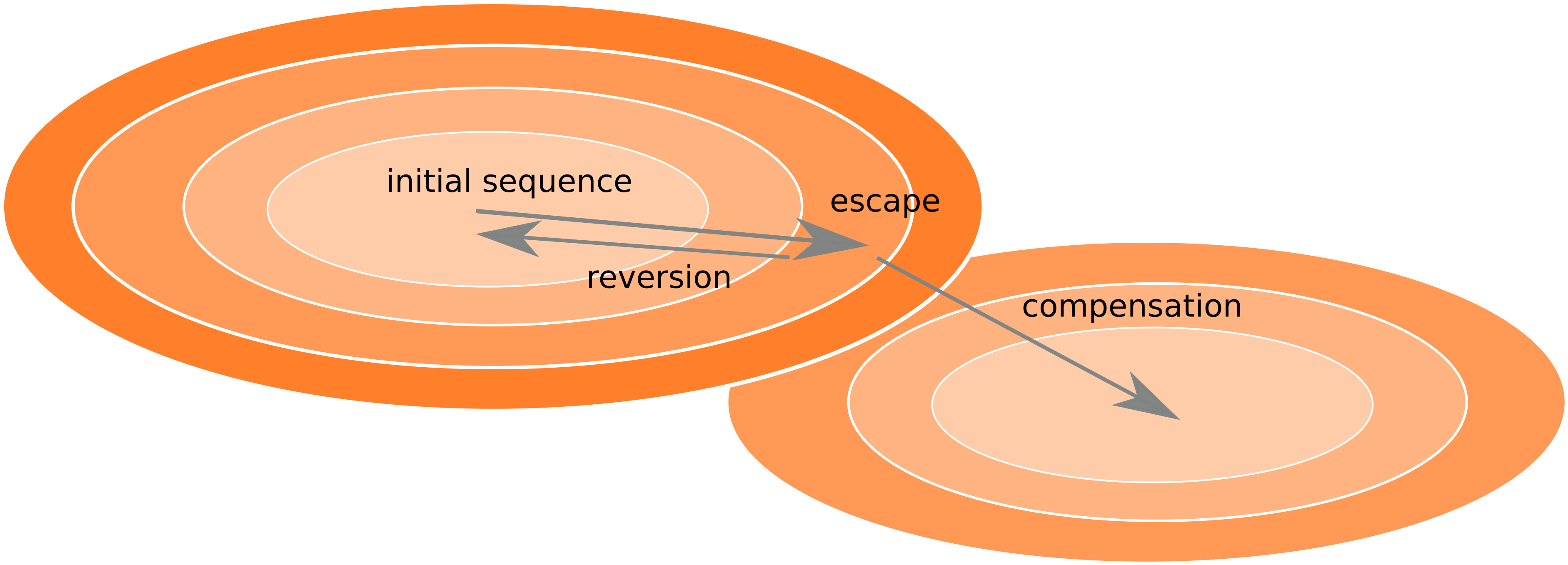
Frequent reversion of previously beneficial mutations
- HIV escapes immune systems
- most mutations are costly
- humans selects for different mutations
- compensation or reversion?
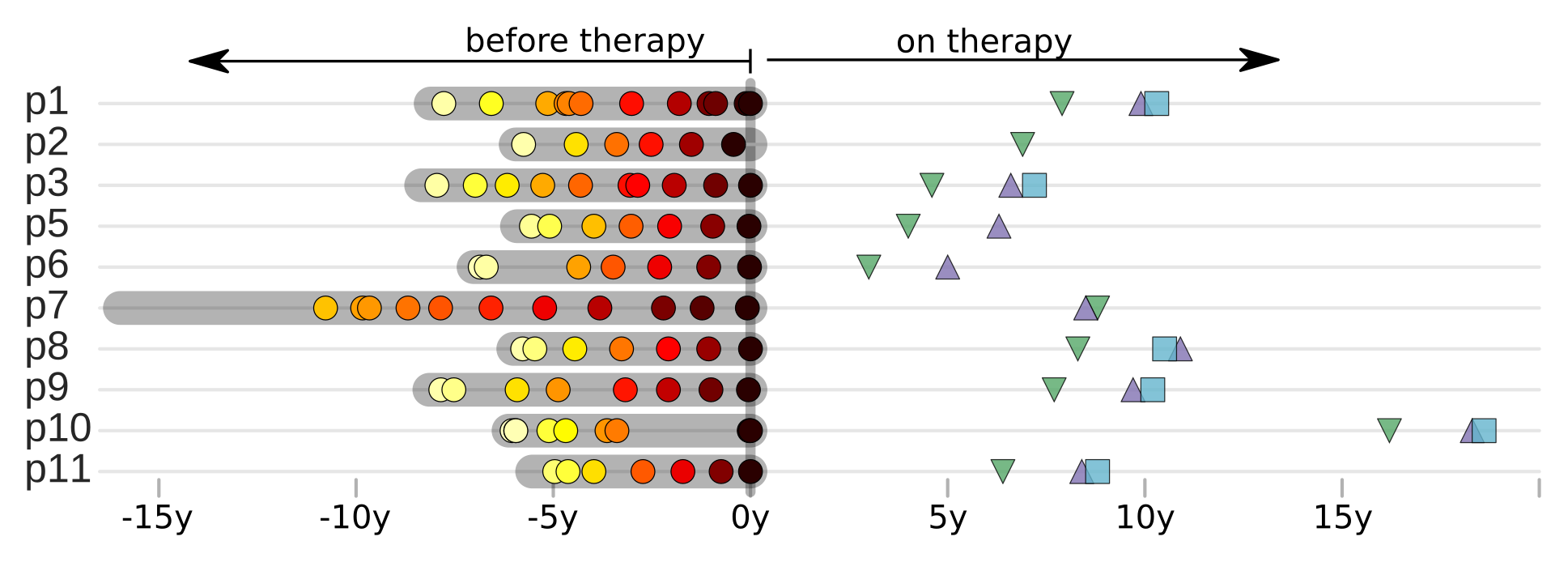
Does HIV evolve during therapy?
Brodin et al, eLife, 2016
No evidence of ongoing replication
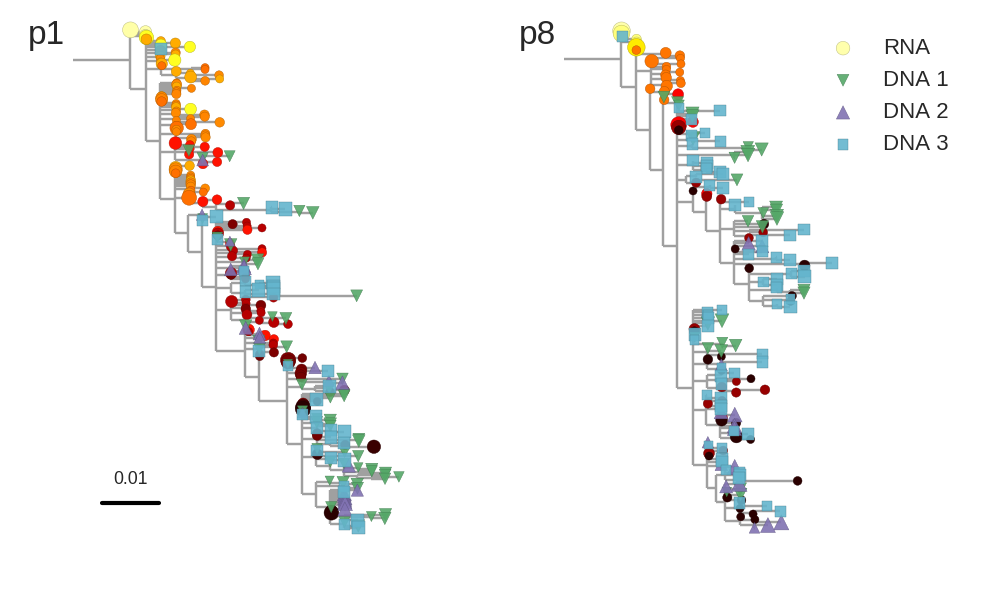
No evidence of ongoing replication
 Brodin et al, eLife, 2016
Brodin et al, eLife, 2016
T-cell turnover is fast in untreated infection

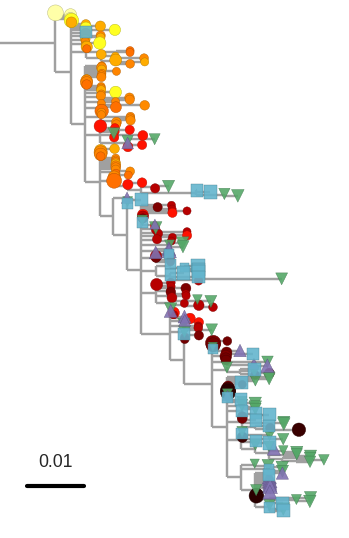
- latent HIV → barcode of a T-cell lineage
- all latent integrated virus derives from late infection
- untreated: T-cell lineages are short lived
- on therapy: T-cell clones live decades
nextstrain.org
joint work with Trevor Bedford & his lab
code at github.com/nextstrain
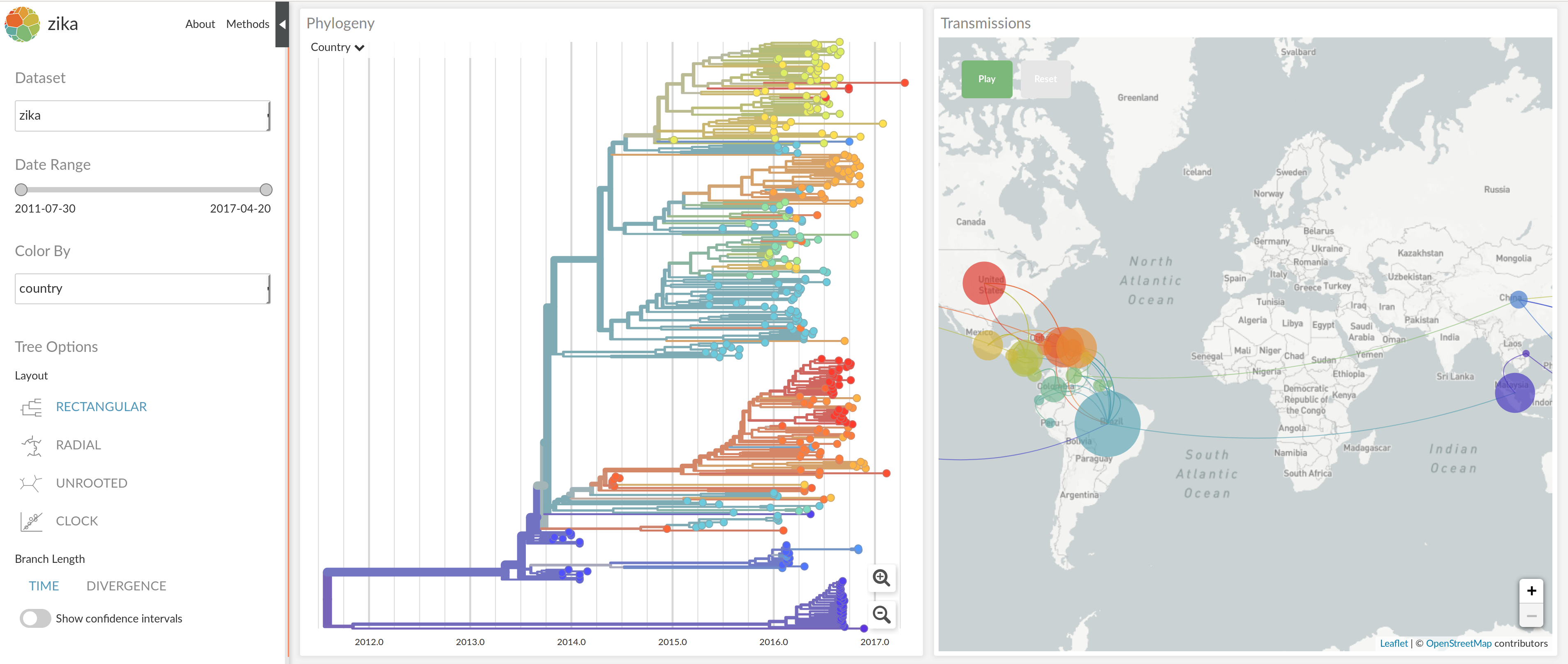
Summary
- Intra-host HIV evolution is governed by a universal fitness landscape, modulated by host-specific immune response
- Landscape of fitness costs can be estimated from intra-host diversity
- The latent HIV DNA reservoir turns over fast
- No evidence for evolution under therapy
- all data are available at hiv.biozentrum.unibas.ch
Thank you for your attention!
Acknowledgments
- Fabio Zanini
- Jan Albert
- Johanna Brodin
- Christa Lanz
- Göran Bratt
- Lina Thebo
- Vadim Puller
nextstrain.org
- Trevor Bedford
- Colin Megill
- Pavel Sagulenko
- Sidney Bell
- James Hadfield
- Wei Ding
- Tom Sibley
- Emma Hodcroft

Amplification bias and template input

Accuracy of minor variant frequencies
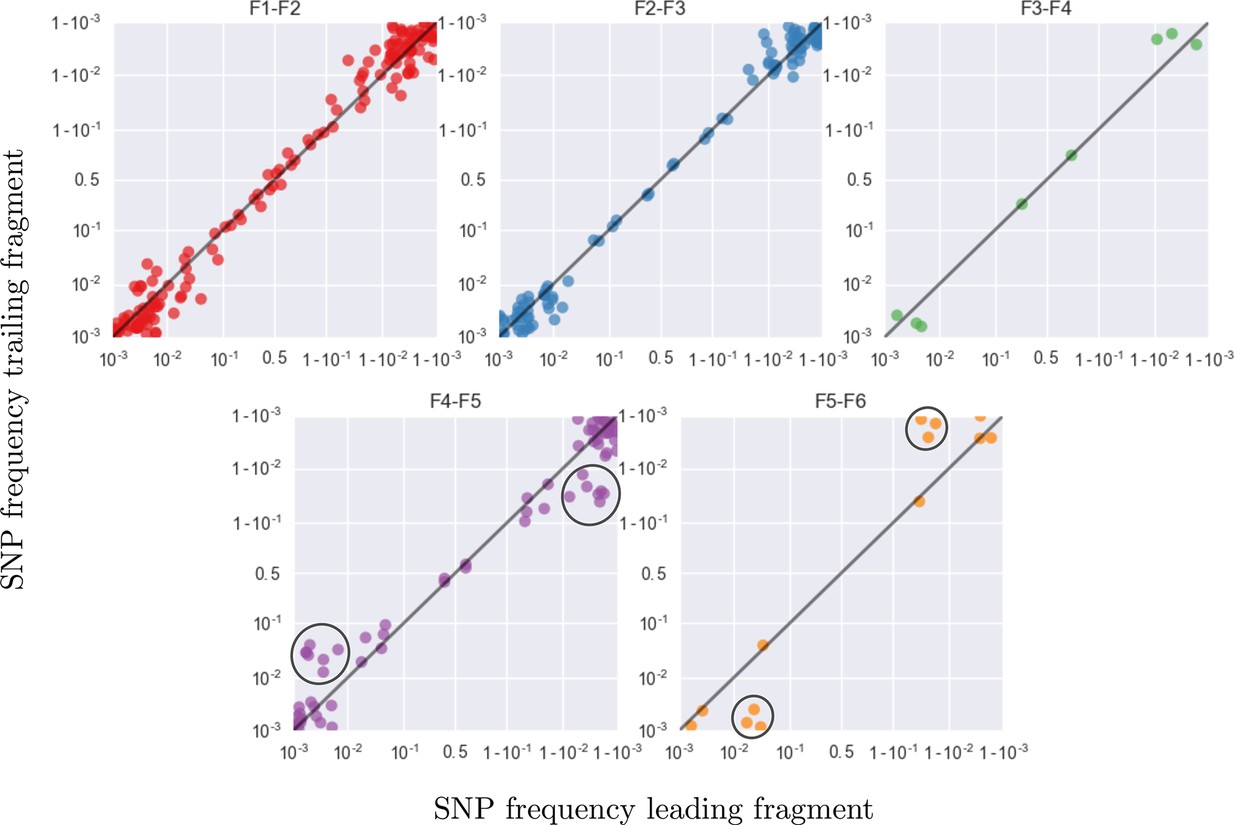
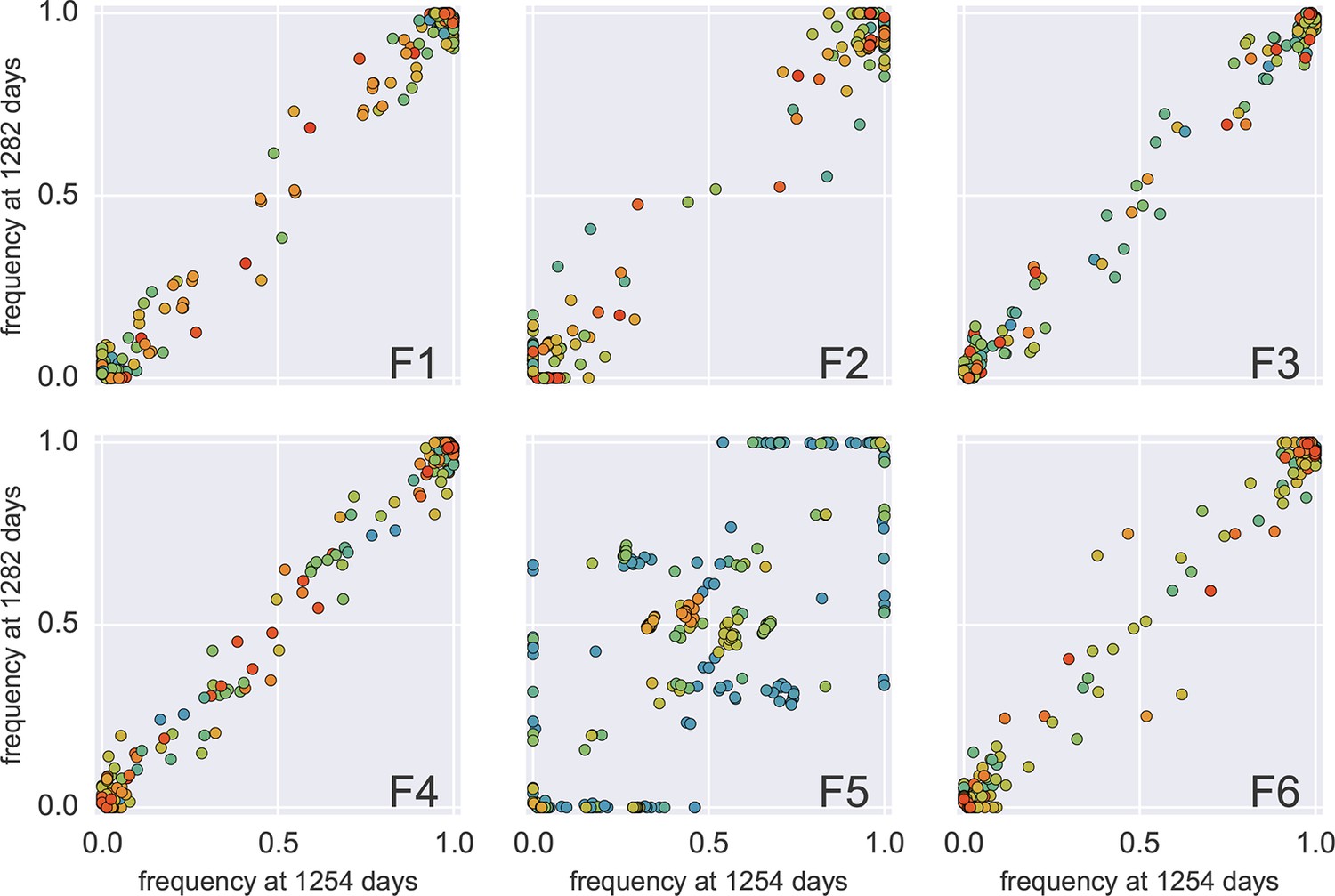
Frequency concordance in samples 4 weeks apart
The distribution of fitness costs
Zanini et al, Virus Evolution, 2017Fitness costs vs consensus amino acid
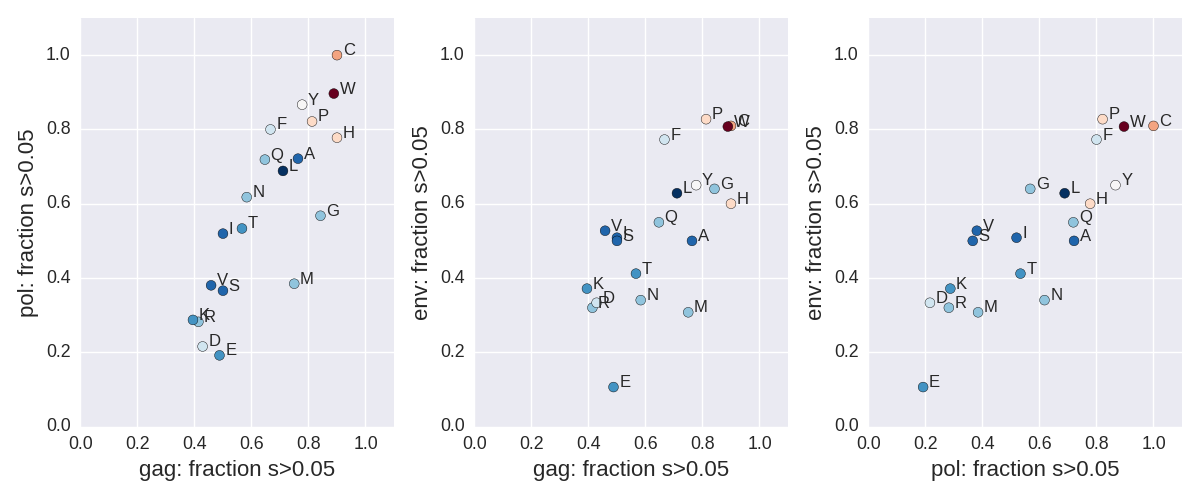 Zanini et al, Virus Evolution, 2017
Zanini et al, Virus Evolution, 2017